Bouhassira Lab publications
Complete list of publications can be found here.
Production of red blood cells
Taking advantage of advancements in the methods to derive human embryonic stem cells and produce induced pluripotent stem cells, we have been working on developing more efficient ways to differentiate pluripotent stem cells into red blood cells since 2004.
This has led to the development of PSC-RED, a highly efficient and cost-effective chemically defined method to produce functional red blood cells in culture from iPSCs.
The ability to produce large numbers of red blood cells in culture will help create new diagnostic tools, and provide cells for transfusions for disorders such as sickle cell disease and myelo-displastic syndrome that often result in allo-immunization due to the need for frequent transfusions.
In vitro produced red blood cells can also be genetically modified to express therapeutic proteins and used as drug delivery vehicles.
- PSC-RED and MNC-RED: Albumin-free and low-transferrin robust erythroid differentiation protocols to produce human enucleated red blood cells. Olivier EN, Zhang S, Yan Z, Suzuka S, Roberts K, Wang K, Bouhassira EE.
Experimental Hematology 2019 Jul;75:31-52.e15; PMID:31176681. Cited by 17
. - Ultra-High-Frequency Reprogramming of Individual Long-Term Hematopoietic Stem Cells Yields Low Somatic Variant Induced Pluripotent Stem Cells. Wang K, Guzman AK, Yan Z, Zhang S, Hu MY, Hamaneh MB, Yu YK, Tolu S, Zhang J, Kanavy HE, Ye K, Bartholdy B, Bouhassira EE.
Cell Reports 2019 Mar 5;26(10):2580-2592.e7 PMID:30840. Cited by 10
- Globin switches in yolk sac primitive and fetal definitive RBCs produced from human embryonic stem cells. Qiu C, Olivier EN, Velho M, Bouhassira EE.
Blood 2008; 111(4):2400-8. PMID:18024790. Cited by 178
- Large-scale production of embryonic red blood cells from human embryonic stem cells. Olivier EN, Qiu C, Velho M, Hirsch RE, Bouhassira EE.
Experimental Hematology 2006 Dec;34(12):1635-42. PMID:17157159. Cited by 180
Gene therapy for the hemoglobinopathies
We have been researching gene therapy for hemoglobinopathies for several years. Our initial research focused on understanding the causes of transgene silencing in erythroid cells. Using the Recombinase Mediated Cassette Exchange method (RMCE), we discovered that silencing in eukaryotic cells is not mainly due to integration near heterochromatin but rather due to transcriptional interferences between the transgenes and neighboring sequences.
By studying expression cassettes without CpG dinucleotides or pre-methylated prior to integration, we determined that DNA methylation is not crucial for silencing but rather confers an epigenetic memory.
We have also observed that the inclusion of insulators in gene therapy cassettes can be both beneficial and harmful. These elements can prevent silencing and insertional mutagenesis at some genomic integration sites, but they can also cause silencing and insertional mutagenesis at other sites.
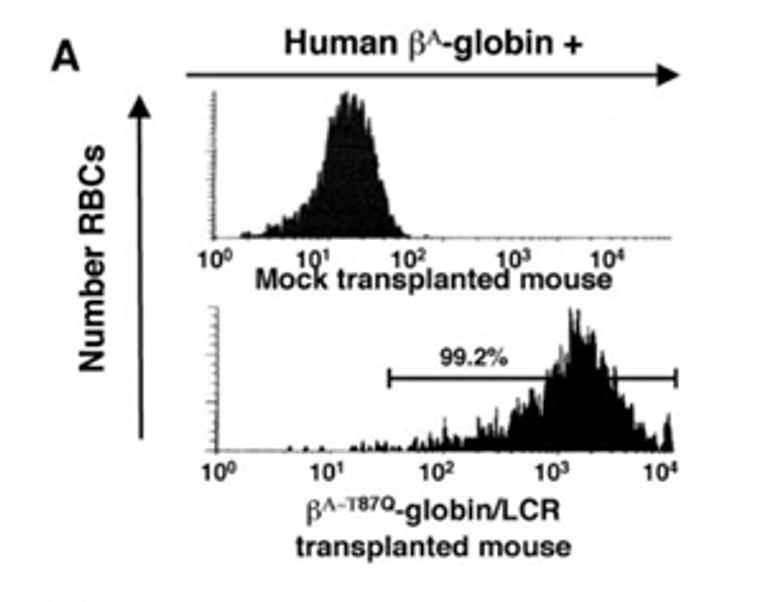
In collaboration with the Leboulch, Humphries, and Eaves labs, we used our findings to design lentiviral vectors, which provided proof of concept that gene therapy could cure sickle cell disease in a mouse model. These vectors were taken to clinical trials by Bluebird Bio and are now FDA-approved for certain gene therapy applications.
We demonstrated in an human iPSC model that it is possible to cure an hemoglobinopathie using a knock in approach.
More recently, we collaborated with the Sadelain lab, who were conducting a gene therapy clinical trial for beta-thalassemia using a reduced conditioning approach.
We are currently testing a novel CRSPR/Cas9-based method to treat the hemoglobinopathies using a knock-in strategy. This method will help treat patients not eligible for existing therapies.
- Lentiviral globin gene therapy with reduced-intensity conditioning in adults with β-thalassemia: a phase 1 trial. Boulad F, Maggio A, Xiuyan Wang X, Moi P, Acuto S, Kogel F, Takpradit A, Prockop S, Mansilla-Soto J, Cabriolu A, Odak A. Thummar K, Du F, Shen L, Raso s, Barone R, Di Maggio R, Pitrolo L, Giambona A, Mingoia M, Everett JK, Hokama P, Roche A, Cantu A, Adhikari H, Reddy S, Bouhassira EE, Mohandas N, Bushman FD, Rivière I, Sadelain M.
Nat Medicine 2022 Jan;28(1):63-70. PMID:34980909. Cited by 10
- Correction of sickle cell disease in transgenic mouse models by gene therapy. Pawliuk R., KA Westerman, ME Fabry, E Payen, R Tighe, EE Bouhassira, SA Acharya, J Ellis, IM London, CJ Eaves, RK Humphries, Y Beuzard, RL Nagel, P Leboulch (2001):
Science. 2001, 294:2368-71. PMID:11743206. Cited by 663
- DNA methylation supports intrinsic epigenetic memory in mammalian cells. Feng YQ, Desprat R, Fu H, Olivier E, Lin CM, Lobell A, Gowda SN, Aladjem MI, Bouhassira EE.
PLoS Genetics 2006 Apr;2(4):e65. PMID:16683039. Cited by 79
- The Human beta-Globin Locus Control Region Can Silence as Well as Activate Gene Expression. Feng YQ, Warin R, Li T, Olivier E, Besse A, Lobell A, Fu H, Lin CM, Aladjem MI, Bouhassira EE. (2005):
Molecular and Cellullar Biology 2005, 25:(10):3864-74. PMID:15870261. Cited by 49
Recombinase-Mediated Cassette Exchange and safe-harbor concept
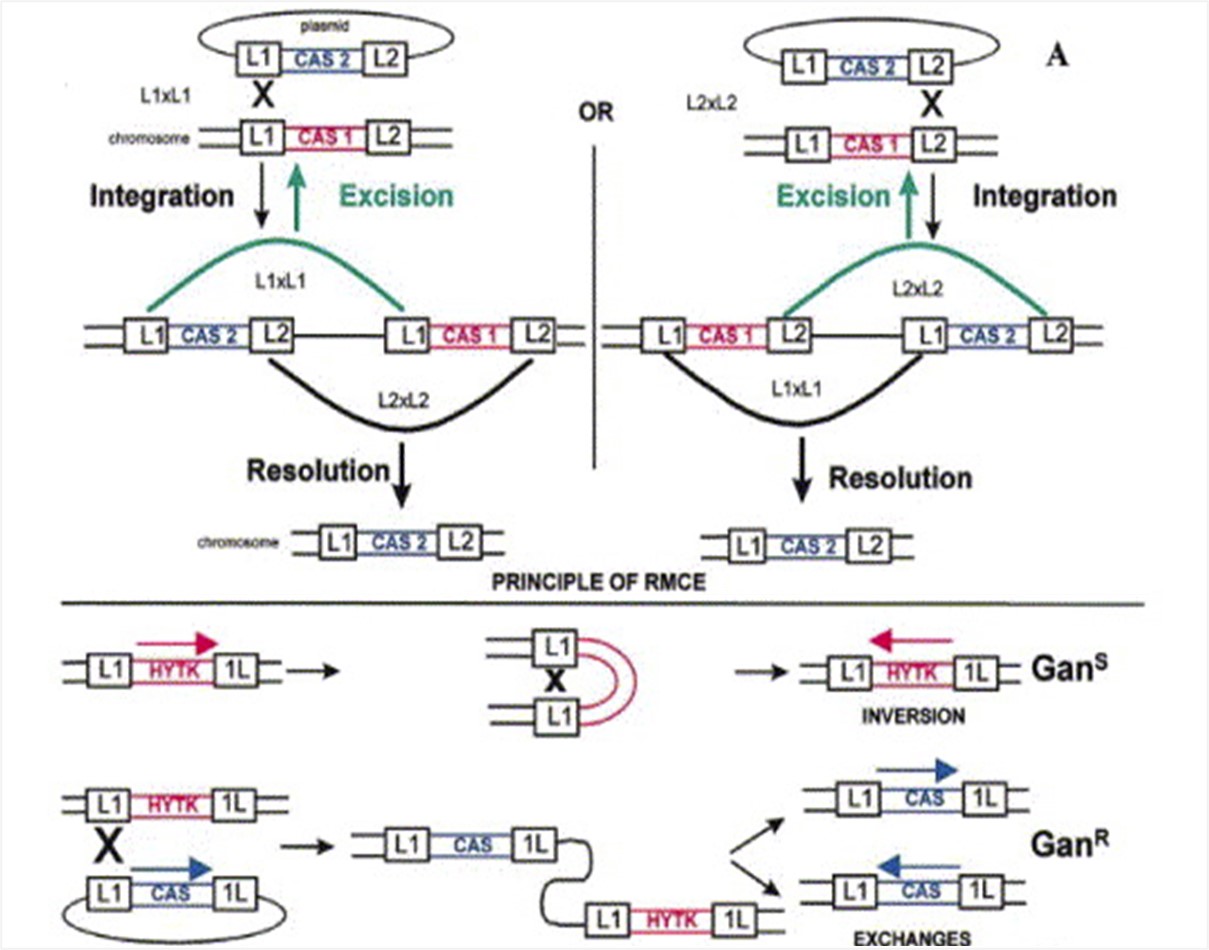
Lentiviral transduction, stable transfection, and transgenesis can result in random integration of transgenes, causing positive or negative position-effects, which complicate gene therapy and genetic engineering. We developped Recombinase Mediated Cassette Exchange (RMCE) to overcome these problems by enabling site-specific integration of cassettes at predetermined chromosomal locations in mammalian cells.
We used RMCE extensively to better understand the molecular basis of position-effects. RMCE has been widely adopted and used in over 800 studies, leading to the development of the concept of safe harbors for gene therapy.
Safe harbors provide a cost-efficient and safe alternative to traditional gene therapy methods, eliminating the need for designing and testing new vectors or gene editing strategies for every defective gene.
We demonstrated the use of safe harbors in a successful cure of alpha-thalassemia in an iPSCs model using a therapeutic construct integrated at the AAVS1 genomic site.
.- Transcriptional Behavior of LCR Elements Integrated At The Same Chromosomal Locus By RMCE. Bouhassira EE., K Westerman, P Leboulch:
Blood 1997, 90: 3332-3244. PMID:9345015. Cited by 195
- Site-Specific Chromosomal Integration In Mammalian cells: Highly Efficient CRE Recombinase-Mediated Cassette Exchange. Feng YQ, Seibler J, Alami R, Eisen A, Fiering SN, Bouhassira EE:
Journal of Molecular Biology 1999 292 (4): 779-785. PMID:10525404. Cited by 278
- Zinc-finger nuclease mediated correction of α-thalassemia in iPS cells. Chang CJ and Bouhassira EE.
Blood 2012; Nov 8;120(19):3906-14. PMID:23002118. Cited by 116
Regulation of replication in erythroid cells
Our studies using the Recombinase-Mediated Cassette Exchange (RMCE) method and globin transgenes revealed that replication timing plays a critical role in gene expression in erythroid cells.
To better understand the relationship between replication and trasnscription, we investigated the mechanisms of replication regulation in basophilic erythroblasts.
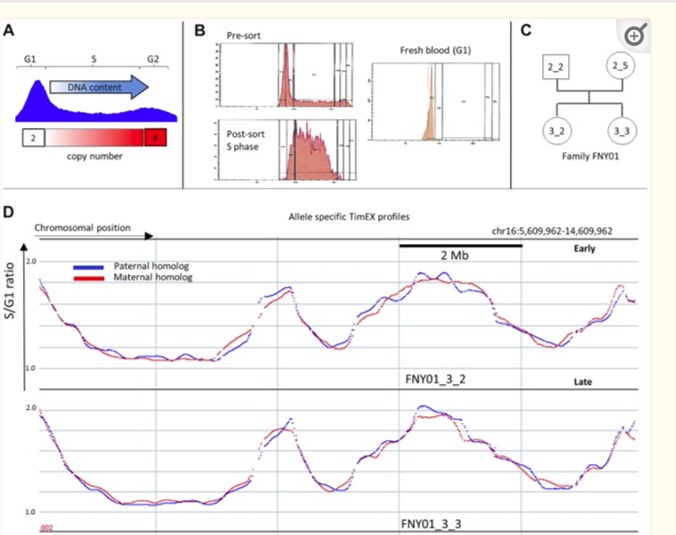
We developed the TimEX-seq methods to measure genome-wide replication timing using massively parallel sequencing. Our results showed that replication timing is tightly regulated in mammalian cells and closely linked to gene expression levels.
We also created methods for generating allele-specific profiles of replication timing and replication origins.
Our analysis revealed that 92% of the genome replicates within a few minutes of each other, while 8% replicates asynchronously, associated with imprinting and proximity to large structural variants.
Finally, we also found that the propensity of origins to unwind and adopt non-B DNA structures, rather than the ability to form G4-quadruplexes, is directly related to origin activity and can be determined by an asymmetry in nucleotide distribution.
- Epigenetic control of replication timing and transcription at hundreds of autosomal loci that are essential for chromosome stability. Heskett, MB, Vouzas AE, Smith LG, Boniface C, Bouhassira EE, Spellman P, and Gilbert DM and Thayer MJ.
Nature Communications 2022, 13,6301
- Allele-specific analysis of DNA replication origins in mammalian cells.Bartholdy B, Mukhopadhyay R, Lajugie J, Aladjem MI, Bouhassira EE.
Nature Communications 2015 May 19;6:7051. PMID:25987481. Cited by 48
- Allele-Specific Genome-wide Profiling in Human Primary Erythroblasts Reveal Replication Program Organization. Mukhopadhyay R, Lajugie J, Fourel N, Selzer A, Schizas M, Bartholdy B, Mar J, Lin CM, Martin MM , Ryan M, Aladjem MI and Bouhassira EE.
PLoS Genetics 2014 May 1;10(5):e1004319. PMID:24787348 Cited by 58
- Predictable dynamic program of timing of DNA replication in human cells. Desprat R, Thierry-Mieg D, Lailler N, Lajugie J, Schildkraut C, Thierry-Mieg J, Bouhassira EE.
Genome Research 2009 Dec;19(12):2288-99. PMID:19767418 Cited by 135
- Human Replicators Can Prevent Gene Silencing. Fu H., Lixin W., Lin CH, Singhania S, Bouhassira EE, Aladjem MI.
Nature Biotechnology 2006, 24(5):572-6. PMID:16604060 Cited by 39
Bioinformatics: GenPlay, the Einstein genome analyzer
Our work on epigenetic led to the creation of GenPlay, a powerful and user-friendly open-source genome browser and analysis tool.
While there are many genome browsers available, GenPlay is unique because of its ability to perform operations on the data, and to display multiple genomes at the same time calcualting the coordinates on the fly.
The project is currently dormant but GenPlay remains one the fastest and funnest Genome Browser around.
The GenPlay genome browser and its full documentation can be accessed here.
- GenPlay Multi-Genome, a tool to compare and analyze multiple human genomes in a graphical interface. Lajugie J, Fourel N, Bouhassira EE.
Bioinformatics 2015 Jan 1;31(1):109-11. PMID:25178461. Cited 7
- GenPlay, a multipurpose genome analyzer and browser. Lajugie J, Bouhassira EE.
Bioinformatics 2011 Jul 15;27(14):1889-93. PMID:21596789. Cited 26
Molecular basis of Thrombotic Thrombocytopenic Purpura (TTP)

In collaboration with Dr.Han-Mou Tsai we have demonstrated that ADAMTS13 is responsible for congenital TTP, that auto-antibody resistant forms of the protein could be generated, that ADMTS13 is expressed predominantly in stellate cells, and we have explored some of the molecular mechanisms associated with low levels of ADAMTS13 expression in human population.
We are currently developing engineered red blood cells carrying a membrane bound fragment of ADAMTS13 that is resistant to auto-anti-ADAMTS13 antibodies, and trying to provide a proof of princple taht transfusion of these cells can be used to treat TTP.
- An IAP retrotransposon in the mouse ADAMTS13 gene creates ADAMTS13 variant proteins that are less effective in cleaving von Willebrand factor multimers. Zhou W, Bouhassira EE, Tsai HM. Lajugie J, Fourel N, Bouhassira EE.
Blood 2007; 110(3):886-93. PMID:17426255. Cited by 43
- ADAMTS13 is expressed in hepatic stellate cells. Laboratory investigation; a journal of technical methods and pathology. Zhou W, Inada M, Lee TP, Benten D, Lyubsky S, Bouhassira EE, Gupta S, Tsai HM.
Laboratory Investigations 2005; 85(6):780-8. PMID: 15806136 Cited by 194
- Enzymatically active ADAMTS13 variants are not inhibited by anti-ADAMTS13 autoantibodies: a novel therapeutic strategy? Zhou W, Dong L, Ginsburg D, Bouhassira EE, Tsai HM.
The Journal of Biological Chemistry 2005; 280(48):39934-41. PMID:16203734. Cited by 59
- Mutations in a member of the ADAMTS gene family cause thrombotic thrombocytopenic purpura. Levy GG, Nichols WC, Lian EC, Foroud T, McClintick JN, McGee BM, Yang AY, Siemieniak DR, Stark KR, Gruppo R, Sarode R, Shurin SB, Chandrasekaran V, Stabler SP, Sabio H, Bouhassira EE, Upshaw JD Jr, Ginsburg D, Tsai HM.
Nature 2001; 413(6855):488-94. PMID:11586351. Cited by 1954
H1 Linker histones
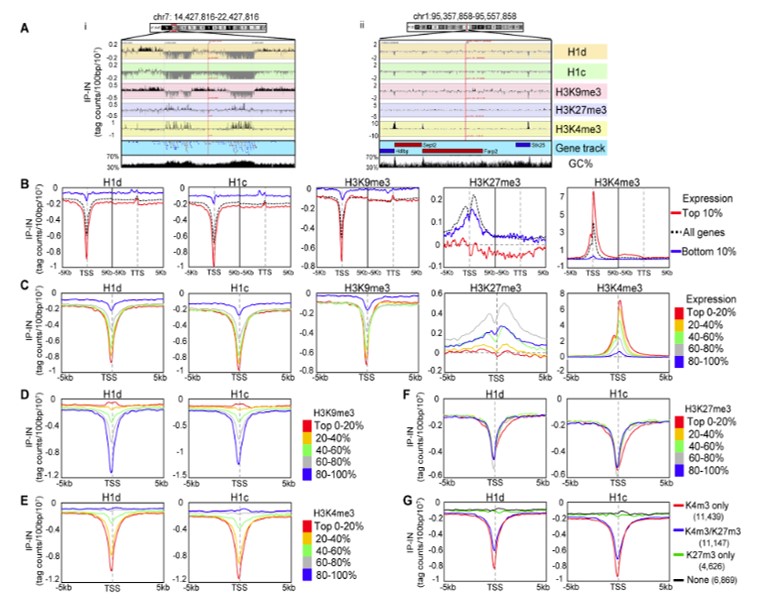
Linker histones are chromatin protein that play essential roles in packing the DNA into the nucleus. They bind the DNA between the nucleosome octamer and are believe to modulate expression and DNA replication
We became interested in linker histones because experiments in which we crossed mice expressing globin transgenes that silenced over time with mice knocked-out for individual H1 linker histones isoforms revealed that the rate of silencing was dependent on the type of linker histone remaining in the cells. This observation led us to collaborate with the Skoultchi and Fan labs to better understand the function of these proteins and their effect on transcription.
These studies led to the observation that while individual linker histone isoforms were largely dispensable, knocking-out three of the seven led to embryonic lethality.
We also produced genome-wide map of the distribution of linker histones using teh CHip-seq methods and demonstrated that linker histone depletion altered the expressionof a subset of imprinted genes, .
-
Histone h1.3 suppresses h19 noncoding RNA expression and cell growth of ovarian cancer cells. Medrzycki M, Zhang Y, Zhang W, Cao K, Pan C, Lailler N, McDonald JF, Bouhassira EE, Fan Y.
Cancer Research 2014 Cancer Res. Nov 15;74(22):6463-73. PMID: 25205099 Cited by 87
- High-resolution mapping of h1 linker histone variants in embryonic stem cells.Cao K, Lailler N, Zhang Y, Kumar A, Uppal K, Liu Z, Lee EK, Wu H, Medrzycki M, Pan C, Ho PY, Cooper GP Jr, Dong X, Bock C, Bouhassira EE, Fan Y.
PLoS Genetics 2013 PLoS Genet. 9(4):e1003417. PMID:23633960 Cited by 115
- Histone H1 depletion in mammals alters global chromatin structure but causes specific changes in gene regulation. Fan Y, Nikitina T, Zhao J, Fleury TJ, Bhattacharyya R, Bouhassira EE, Stein A, Woodcock CL, Skoultchi AI.
Cell 2005 Cell. Dec 29;123(7):1199-212 PMID:16377562 cited by 161
- Mammalian linker-histone subtypes differentially affect gene expression in vivo. Alami R, Fan Y, Pack S, Sonbuchner TM, Besse A, Lin Q, Greally JM, Skoultchi AI, Bouhassira EE.
Proc Natl Acad Sci U S A. 2003 May 13;100(10):5920-5. PMID:12719535